
公共卫生学术热点追踪
Nature | 王开拓等解析丙肝病毒HCV表面糖蛋白E1/E2高阶复合物结构
慢性丙型肝炎(Chronic Hepatitis C)是由丙肝病毒(Hepatitis C Virus, HCV)引起的一种肝脏炎症。全球约有5000万慢性丙肝病毒感染者,每年约新增100万感染病例。据世界卫生组织估计,2022年约有242,000人死于由丙型肝炎引发的肝硬化和肝癌等并发症。尽管目前已有针对丙肝的特效药物,如索磷布韦、达拉他韦等,但药物价格昂贵,且治愈患者仍有再次感染病毒的风险。因此,推动预防性HCV疫苗的发展具有重大的现实意义。
HCV是一种包膜的单链正链RNA病毒,属于黄病毒科丙型肝炎病毒属。其全长RNA约有9500个碱基对,编码一个超过3000个氨基酸的多聚蛋白前体。该前体在宿主细胞和病毒自身蛋白酶的作用下,裂解成多种独立的病毒蛋白。其中,表面糖蛋白E1和E2位于病毒包膜上,是高度糖基化的蛋白,也是病毒颗粒表面唯一的蛋白质组分。E1和E2以异源二聚体的形式存在于病毒颗粒包膜表面,对病毒侵入细胞至关重要,是现有丙肝病毒中和抗体的主要靶点,也是潜在疫苗开发的主要目标,因此其结构研究受到了广泛关注。然而,E1/E2复合物高度糖基化且结构极不稳定,使得结构解析极具挑战性。
此前,最完整的E1/E2结构研究是在2022年10月发表在Science杂志上的E1/E2与三种单克隆抗体形成的复合物[1]。该研究揭示了E1/E2的异二聚化界面、蛋白表面糖基化修饰以及抗体结合表位等关键结构信息,但复合物的跨膜结构域由于过于动态而未能解析。此外,HCV病毒颗粒表面的E1/E2蛋白被认为会形成更高阶的复合物组装形式。尽管其他包膜病毒如HIV、新冠病毒、流感病毒的表面刺突蛋白常形成三聚体结构,但HCV的E1/E2是否形成高阶聚体一直未得到结构信息的验证[2]。
2024年9月4日,哥本哈根大学免疫学和微生物学系(ISIM)的Jannick Prentø和哥本哈根大学生物医学科学学院的Pontus Gourdon和中国科学院植物研究所王开拓合作在Nature杂志上发表了The hepatitis C virus envelope protein complex is a homodimer of heterodimers。这项工作解析了首个丙肝病毒HCV表面糖蛋白E1/E2的高阶复合物冷冻电镜结构。研究结果表明,体外纯化的HCV E1/E2形成了二聚体组装形式,而非其他包膜病毒常见的三聚体结构。这一发现颠覆了先前的认知,对设计有效的HCV疫苗具有重要的指导意义。

技术上,由病毒学家和结构生物学家组成的联合研究团队在多年的合作研究中进行了多项技术创新,最终取得了这一成果。E1和E2蛋白的N末端和C末端对蛋白结构和功能至关重要,因此无法使用传统标签方法进行纯化。在先前的Science文章研究中,研究者采用带有Strep标签的单克隆抗体与蛋白复合物共表达,以抗原-抗体复合物的形式纯化整个蛋白复合物。而在本研究中,研究者将Strep标签设计在E2蛋白中间的一个柔性loop区,并进行了功能验证,使得单独纯化蛋白复合物成为可能。此外,由于蛋白复合物表达量极低且在体外体系中极不稳定,研究者克服了蛋白纯化和冷冻制样过程中的多项技术难题。在每次大规模纯化中仅能得到小于10微克样品的情况下,研究者成功对蛋白进行了结构解析。
针对蛋白样品的柔性,研究者们采用了多种新的冷冻电镜结构解析技术、密度加强及模型搭建算法。特别是在Alphafold[3]结构预测和基于深度学习的三维密度图后处理算法EMReady[4]的帮助下,研究者构建了几乎所有氨基酸序列的结构模型,覆盖了E1序列的96%和E2序列的90%。相比之下,Science文章中的结构模型仅包括51%的E1和82%的E2序列。
文章中最重要的发现是E1/E2异聚体能进一步组装成更高阶的二聚体复合物。这种二聚化是由E2蛋白介导的,相互作用面积约为1200平方埃,主要基于位于蛋白表面的多个氨基酸间的疏水或氢键相互作用,而蛋白表面的糖基基本不参与二聚化。二聚化使得此区域被包裹在复合体内部,而其他暴露区域表面多数都是高度糖基化的,这可能解释了HCV E1/E2糖蛋白复合物的低免疫原性。作者在此基础上开展了一系列的蛋白突变体生化和免疫实验,证明了二聚体界面中的氨基酸序列非常保守,对其突变会影响二聚体的稳定性。此外,先前的研究结果也显示,位于二聚化界面上至少一半的氨基酸残基在所有亚型HCV中都完全保守,且其突变会影响HCV假病毒颗粒(HCV pseudoparticle,或者HCVpp)对细胞的侵染活性[5]。
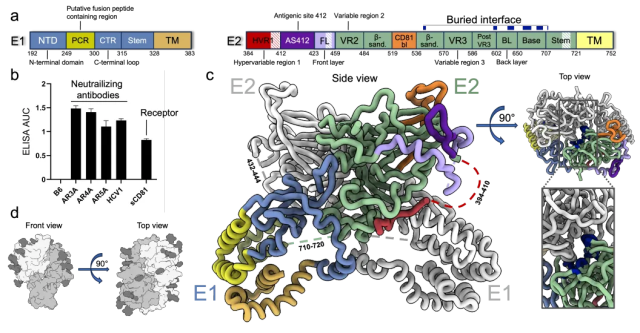
图二:HCV E1/E2蛋白复合物的冷冻电镜结构。
此次解析的结构还揭示了多种此前研究中未被观察到的结构细节。首先,复合物的跨膜螺旋区结构首次得到解析,复合物结构中的跨膜蛋白束排列较松散,平均分辨率约5埃,但结合Alphafold结构预测,研究者仍能搭建来自两个蛋白的总共6个跨膜螺旋模型。其中,M1-M2来自E1蛋白的PCR (putative fusion peptide containing region)区域,M1和M2螺旋中间的链接区域(272-281位氨基酸)非常保守,被认为是介导病毒颗粒与侵染细胞融合的肽段(putative fusion peptide,简称pFP);M3-M4来自E1蛋白的C端,M3前面还发现了一个可能平行于膜所在平面的螺旋,命名为MX螺旋;M5-M6位于E2蛋白的C端。

图三:HCV E1/E2复合物中跨膜结构域的位置和结构。
其次,结构中首次解析了E2蛋白N末端的区域,包括序列最多样化的hypervariable region 1 (HVR1)以及序列保守但结构非常不稳定的antigenic site 412 (AS412)。这些区域被认为对HCV的免疫逃逸至关重要,但其具体作用机制和结构尚不完全清楚。结构分析发现,HVR1区域可以隐藏多种HCV中和抗体的识别表位以及受体CD81的结合表位,而体外实验也证明,样品与多种中和抗体和受体CD81的结合能力在4摄氏度时较低,在温度升高时会升高,这暗示了此部分结构在病毒侵染和被免疫系统识别时需要进行一定程度的重排。
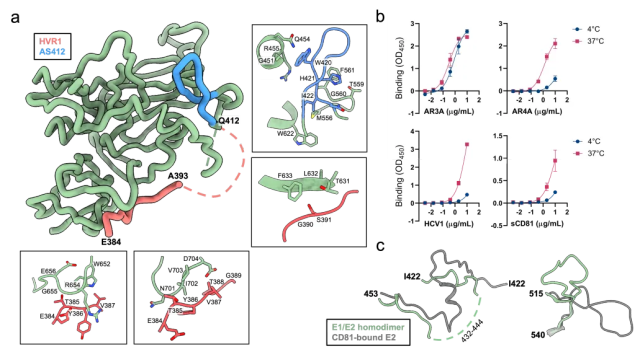
图四:E2蛋白N末端HCV1和AS412区域的结构及其对蛋白结合能力的影响。
文章所揭示的HCV E1/E2高阶结构对于设计有效的HCV疫苗具有重要的指导意义,也是后续结构解析工作的起点。文章的通讯作者,来自哥本哈根大学免疫学和微生物学系(ISIM)的Jannick Prentø和哥本哈根大学生物医学科学学院的Pontus Gourdon,将在此成果的基础上继续开展系统性研究,攻克病毒颗粒中的E1/E2原位结构以及E1/E2介导病毒融合过程不同状态复合物的结构,并在此基础上进行疫苗设计和改造。
(来源:BioArt)
参考文献:
1. Torrents de la Peña A, Sliepen K, Eshun-Wilson L, et al. Structure of the hepatitis C virus E1E2 glycoprotein complex. Science. 2022;378(6617):263-269.
2. Freedman H, Logan MR, Hockman D, Koehler Leman J, Law JLM, Houghton M. Computational Prediction of the Heterodimeric and Higher-Order Structure of gpE1/gpE2 Envelope Glycoproteins Encoded by Hepatitis C Virus. J Virol. 2017;91(8):e02309-16.
3. Jumper J, Evans R, Pritzel A, et al. Highly accurate protein structure prediction with AlphaFold. Nature. 2021;596(7873):583-589.
4. He J, Li T, Huang SY. Improvement of cryo-EM maps by simultaneous local and non-local deep learning. Nat Commun. 2023;14(1):3217.
5. Pfaff-Kilgore JM, Davidson E, Kadash-Edmondson K, et al. Sites of vulnerability in HCV E1E2 identified by comprehensive functional screening. Cell Rep. 2022;39(8):110859.